 Nanotubules acting as guides in the cell nucleus.
Nanotubules acting as guides in the cell nucleus.
Last update June 1, 2001
page in progress
First, I'll put on some explanations or if you like opinions about the basics of the various areas. I'll not try to compete with complete (thick) textbooks about Cell biology, Biochemistry, and similar areas, though I went over a few of them, even thouroughly enough for some subjects, but give an impression of the knowledge I've found relevant some 5 years ago and still find essential, about cells, cell boundaries, membranes, of course the general knowledge about neurons and their interactions, and some about nerve strains and signal progression over them.
From this point, a lot of science has come into being, about the internals of cells, what goes on in the cytoplasm, what various mechanisms are for neuron firing, (chemically induced, voltage) potentials around the cell membrane, permeability control of membranes, and channels with permeability control through voltage steering, the role of microfilaments and various chemicals in the cytoplasm as catalists and (chemical) reaction initiators.
Recently, lets say for a few years, medical articles appear in various international medical magazines about the application of these basic principles for understanding or interfearing with neoronic behaviour in existing brain tissue, for instance in fruit flies, rats, pigeons (see below), and even human beings. Some of them in lets say traditional medication sense where certain substances are shown to spawn or generate certain neuronal responses, fight certain deseases or gain insight in globally or medium scale oriented mechanisms. Also, there are more than a few magazines and articles discussing the finer modes of operation and effects and basic mechanisms of various brain sections and neurons, up to the point of getting a lot closer to good enough models of neuronic behaviour to take serious at biological (cell) and chemical levels to consider taking such a model as a starting point for sensible computer simulation and analysis.
Nanotubules are of interest in various ways, just like various other particles floating around in the cell nucleus (assuming eukariotic cells), such as congomerates of tubules and even 'buckyballs', lets say diamond basic blocks, regular carbon structures. I was first aware of such half a decade or longer ago by stumbling on the book 'Ultimate computing' by (stuart?) Hammeroff, having looked for books on computing rather than the brain, luckily. I did have an interest in parallel networks possibly resembling neuronal behaviour, and of course my general knowledge in the area of electrical engineering quite included neural networks (what is a network theorist supposed to know), I even talked with some scientists with working results in the fields, and was all to aware of their very limited capabilities of modeling actual brain behaviour, so the book made me quite aware that on top of the things I already looked for to model neuron behaviour more accurately, there was already more known about what is in them than was needed to raise more interest. Evident questions are: how do axons and dendrites grow, and how is the firing behaviour influenced, how could possible external influences be included in the behaviour picture, etc.
The effect of nanotubules and their raison d'etre in various cells is studied, and I'll mention some articles I found, and I suspect that at least it is not unreasonable to assume that the valid medical field of morphogenetics is not unrelated to their role for instance in mytosis (cell division), their role as guide for transfering vesicles and possibly of an other interesting microbiological phenomenon: motor molecules. These molecules, not completely unlike though not directly related to the idea of spermatozoids propelling themselves though wiggling their tails, are like little chemically powered motores rotating their faglellum tails of a long, flexible molecule, also to propel themselves in the cell fluid they are in. It is not unlikely that nanotubules and nanofilaments are a main factor in the growth of for instance dendrites, acting as rods along which transport and growth take place. The fact that microtubules have interesting and chemical an mechanical properties, such as their cylindrical shape, and for instance the idea that the tubulin molecules in the tube can be in two states, mechanincally, and have even been ascribed pattern moving (counting ?) behaviour makes one wonder what more there can be found in this interesting area.
Evidently a major area directly relates is the vast area of genetics and related fields, which I'll not try to cover. I had a glance recently at an overview of the human genetic map, which is reminding of a major chip design plot of at least A0, which is probably fun enough for genetiticists to have an idea of the progress of a lot of analysises going on all over the world with lots of funding and probably quite a lot at stake, but I can't make chocolate of such research. I lack the knowledge about the various chemicals that make up the dna stands and their behaviour in general, I don't know too much about the known science in the area of recombination, and the many, many reactions and patterns that are known in the area. Also they are not my main interest, which is a pleasant combination of facts, though I want to immedeately add that I am very aware of the fact that the whole process of mytosis (cell division) very much including both the effect of and mechanisms involved in copying of the dna is governed at least in major ways by nano-biological considerations, more specifically by microtubulee aligning in sort of magnetic field patterns directly influencing the cell the division and its results at chemical and biological level.
This has my interest, also at the chemical and physical level, because I'm sure it is essential as a mechanism, up to the point of probably having direct applications in for instance cancer research.
Being quite edified in physics, electronics, and electrical engineering, I was aware of the interest in nano strucures, also in the form of various materials arising as well in biology quite some years ago, and before the current research in this field boomed pretty much. There are quite some industrial and university research activities going on in the fields of micro machining though electronical devices on the nano scale, for instance using few atoms large or wide wires and electronics connections and structures. Making 3d designs of electronics at near atomic scale evidently is of great interest, also for me, and I am aware of various research lines going on there, though this page is not intended to give much introduction or information about these fields, which are more research than development, but still vast and not formed into lets say many working machines. My interest in brain structures, mechanism and behaviour supercedes the interest in those areas lets say on the knowledge level, though I could imagine it would be great fun making some nano devices and let them work.
Mainly, I want to make clear what behaviour and mechanisms are known in the more advanced fields of brain and cell research, including the nano scale and the necessary chemics and physics, and arrive at solid enough starting points to make theories about the possible behaviour of brain cells and conglomerates of them that are solidly rooted in actual medical and other knowledge, and that might be able to get to serious enough simulation models, both mathematically and in the form of computer programs.
I have the idea of making links for the italicized words, for easy reference, as I find time.
I think some of them may not be generally available because the medical library I read them in is subscribed to them, so some links may not work. Various links are links with cgi information in them that may not apply to all browsing persons, so I'm not completely sure they'll work at any point in the future for every person, though I did try myself at some different stations, and they seem fine.
The idea of potential, chemical, and structural energy levels, the concept of heat and entropy.
A short note on quantum mechanics and their fundamental role in determining energy levels and in 'noise', tunneling and probability computations.
Chemical bond types and their general effects, also near the (polar) lipids in the membrane
Energy and electrical potential and current maintenance, pH levels and their barrier effect.
Opening and closing of various types of channels under chemical and current control
Modeling of such, and of general diffusion, mathematical technique/simple model description
The time ranges are not hard to measure, usually millisecond scales suffice.
Complication level in the formation, replication and effects during mitosis of genetic materials, short overview.
The reason for not going into this field is the complexity involved, the amount of knowledge required to seriously master some specialisms, and the inappropriateness for the intended subjects here. Which evidently is no generalistic scientific observation or statement, because more care and articulation whould be in order to consider such, and there is no reason not to ascribe significant relevance to the subject.
Since this page is in builtup, I'll add more elaborate explanations and accurate pictures later.
Nanotubules acting as guides in the cell nucleus.
 Microbodies in the cytoplasm.
Microbodies in the cytoplasm.
 Nucleus during cell division (notice the role of the various guiding strands and filaments).
Nucleus during cell division (notice the role of the various guiding strands and filaments).
 Mitosis in rat kangaroo cell.
Mitosis in rat kangaroo cell.
 Motility Migration of Organells.
Motility Migration of Organells.
 Motility Flagellar Motility - Red Sulphur Bacterium.
Motility Flagellar Motility - Red Sulphur Bacterium.
"Communication and Concurrency", Robin Milner, Prentice Hall International, 1989
Evidently, one would have to suffer from severe delusions to make such a sort of quest in life amoung other things, and think one can cover those major themes and subjects even reasonably in a normal world. Some of these subjects receive world wide attention, and are therefore the shared focus of many, many scientists, amoung which without any question some brilliant ones, so an individual may chose to engage in the knowledge accumulation scene, possible active, get handy, and maybe develop enough daringness and application of (preferably ones own) intelligence to have a gamble with the larger potential theories, and maybe live to be still taken serious by knowledgeable persons in the area.
Clearly the area of genetics has been receiving a lot of attention for obvious reasons, and requires and is accumulating vast areas of expertise probably few only can seriously oversee. The longer range targets may not always be clear to everyone, the classification or lets say finding of the structure and hopefully meaning of every part of mainly the human dna is of course one of the goals having started massive amounts of work. The second part of the question is an in my opinion almost unoverseeable amount of work to deal with, the former, the lets say structure analysis, strickly as a (bio) chemical job, lets say, seems to get along with massive progress through very major investments. Without daring to make an accurate estimate, I would dare argue that such job alone is not a holy grail of research, once completed more insight is not necessarily gained if no other research or work is using the results, of which genetically based medication and genetic engineering probably form the opposite ends of the spectrum.
My personal interest is mainly in two areas, the global functioning of the brain, and possible conclusions for its communication capabilities, combined with preferably as accurate as possible small scale models of its constituents.
As electrical engineer, I'm aware of how computers can be put to effective and succesfull use as simulator of complicated behaviour, lets say like in electronics circuits and computer circuits, as well as more physically oriented models.
The idea of making accurate models is not enough to destil the behaviour of a whole system, and already a computer contains a limited set of different types of parts which are hard to understand when presented as a network of interconnected parts, even for computer builders, and even when the parts themselves are well defined and represented. Let alone when the parts are unknown, and the whole architecture of the computer is of unknown kind.
And finally, let alone when the computer is of high intriticate complexity, of analog, threedimensional parts, and of a connectivity of the order of 10 billion or so parts with up to easily a hundred interconnections per part, which makes for incredible amounts of structural possibilities.
Yet for a computer designer, most computer designs can be understood within a short enough time when they are of a known kind, and then the result normally is that a good designer good do a fair job of making such a machine themselves, with a good match with the example machine, and the amount of information in terms of bytes for instance to describe such an experiment is overseeable, depending a bit on the type of modeling used. There are several types of parts, interconnections with many repetitions, known standard blocks, and the structure is pretty well categorizable and known by lets say the better designers.
The whole network of transisors for modern computers can easily span tens or hundreds of millions of transistors and wires in between then, which as circuit diagram would be worse then the largest roadmap, and far more complex to read or understand, even for all but maybe very few experts, and even they would find it quite hard to even get a bit of an idea of what an unknown circuit actually would do when only the network of transistors is shown, with not much ordering, or levels of abstraction and sub circuit specifications.
Already in computers, there are quite large numbers of small circuit possibilities, easily dozens or hundreds of different kinds, all with very clear and well defined, differing behaviour, and one would have to master a significant number of them to have some understanding of even an old computer system.
The second article has as subject the subject I was drawn to also quite some years ago: pigeons. Not that it in any direct sense even refers to the direction finding talents of the gall blatter-less birds (as I seem to remember), but it does mention the interesting idea of 'fire rate', which I sort of introduced in my thinking and some simple communications years ago stil at Delft University, thinking about possible mechanism for measuring external (field) influences on the brain possibly explaining the pigeon direction sense and or possibly the brain-brain communication mechanisms.
Funny enough, both articles are neither from the us nor form europe, for whatever such formalistic or very meta observation is worth in these days.
from: Neuroscience, Pergamon, vol 102 num 1, 2d jan 2001
"Persistent sodium channel activity mediates subthreshold membrane
potential oscillations and low-threshold spikes in rat entorhinal cortex
layer V neurons"
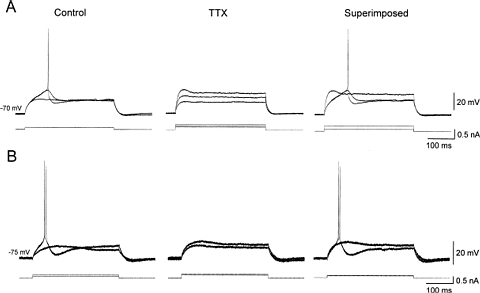
Neurons from layer V of the EC exhibit a low-threshold depolarizing response. (A, B) The left panels (Control) illustrate the responses elicited in two different layer V cells by depolarizing current pulses applied from a holding potential of -70 mV (A) or -75 mV (B). Note in the cell in A that a 0.1-nA depolarizing current pulse resulted in a passive response (lower trace), while a slightly larger pulse (0.11 nA; amplitude change not distinguishable in the figure) triggered a slow regenerative potential (upper traces) that could culminate with a fast action potential. Similarly, in the cell in B, a 0.1-nA depolarizing current pulse triggers a passive response, while a 0.2-nA pulse elicits a slow regenerative response that culminates with a spike doublet. The traces in the middle panels (TTX) demonstrate that in both cells the slow low-threshold regenerative responses were fully abolished by TTX (1 M). In the right panels (Superimposed), the largest amplitude traces from control (left) and TTX (middle) have been laid on top of each other.
Clear resemblance of the graph shape with the ADSR response of the familiar synth envelope generator.
Abstract:
Abstract
Entorhinal cortex layer V occupies a critical position in temporal lobe circuitry since, on the one hand, it serves as the main conduit for the flow of information out of the hippocampal formation back to the neocortex and, on the other, it closes a hippocampal�entorhinal loop by projecting upon the superficial cell layers that give rise to the perforant path. Recent in vitro electrophysiological studies have shown that rat entorhinal cortex layer V cells are endowed with the ability to generate subthreshold oscillations and all-or-none, low-threshold depolarizing potentials. In the present study, by applying current-clamp, voltage-clamp and single-channel recording techniques in rat slices and dissociated neurons, we investigated whether entorhinal cortex layer V cells express a persistent sodium current and sustained sodium channel activity to evaluate the contribution of this activity to the subthreshold behavior of the cells. Sharp-electrode recording in slices demonstrated that layer V cells display tetrodotoxin-sensitive inward rectification in the depolarizing direction, suggesting that a persistent sodium current is present in the cells. Subthreshold oscillations and low-threshold regenerative events were also abolished by tetrodotoxin, suggesting that their generation also requires the activation of such a low-threshold sodium current. The presence of a persistent sodium current was confirmed in whole-cell voltage-clamp experiments, which revealed that its activation "threshold" was negative by about 10 mV to that of the transient sodium current. Furthermore, stationary noise analysis and cell-attached, patch-clamp recordings indicated that whole-cell persistent sodium currents were mediated by persistent sodium channel activity, consisting of relatively high-conductance (~18 pS) sustained openings.
The presence of a persistent sodium current in entorhinal cortex layer V cells can cause the generation of oscillatory behavior, bursting activity and sustained discharge; this might be implicated in the encoding of memories in which the entorhinal cortex participates but, under pathological situations, may also contribute to epileptogenesis and neurodegeneration.
Author Keywords: oscillations; sodium current; parahippocampal; rhythms; low-threshold spike; epilepsy
Abbreviations: EC, entorhinal cortex; EGTA, ethyleneglycolbis(aminoethyl ether)tetra-acetate; HEPES, N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid; INa, transient sodium current; INaP, persistent sodium current; INaW, window sodium current; IR, inward rectification; PIPES, piperazine-N,N'-bis-2-ethanesulfonic acid; TEA, tetraethylammonium; TTX, tetrodotoxin
What does this all mean and refer to?
In short, a rat's brain cell is taken and examined for its
behaviour, and the article makes clear there are additional
mechanisms or parameters to the lets say well known enough,
established theory about neuronal functioning.
Neurons are the main atomic constituents of the brain, the brain is made of neurons connected with eachother through axons and dendrites which run like many irregularly shaped wires between between them. When a neuron 'fires', the synapses between the two carry a mainly chemical signal, and when that signal is generated at the axons of a neuron that fires as a response, it may fire connected neurons in turn, possibly leading to various firing patterns unrolling, maybe terminating at some point, maybe looping and repeating at others.
...
The article in Neuroscience, Pergamon.
This article mentions the fireing rate of neurons, which is used as measurment to compate neuronal responses to various visual patterns being input.
One should be aware of the idea that I'm not at all novice to computer simulation, I've built computer model programs myself that worked for subjects at least ranging from electronics crcuit simulations through 3D modeling and light simulation, and used (also professionally) quite some others.
At this point this text will contain not too much detail on background knowledge I tend or intend to apply, I think some more pages could be in order for that purpose, which is quite a good and usefull one, though inherently complicated, lets say it is a major subject in a serious toplevel electrical engineering curriculum.
At this point, I'll pay some attention to lets say experience models mentioned in various sources I found, mainly based on (chemical) reaction equations coupled with mechanincs and possibly physics and some electrical considerations, leading to (from what I found thus far) diffusion, transport and streaming differential equations, partial differetial equations, application of stokes theorem maybe, and averaging integrals and statistics.
An example of a computer simulation is presented in the virtual cell pages, where a java applet links with a computer with simulator program to do the cell behaviour simulation, at at least advanced and serious level, though not including neuronic behaviour (for one cell) as far as I saw at this point.
The applet requires a (free and easy) registration to use, and java 1.3 (free from e.g. the Sun site), and there are manual pages available. As an example, here is one differential equation as part of a cell model from the applet:
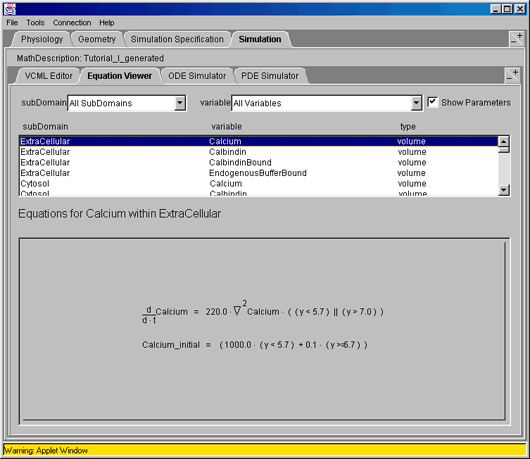
for Virtual Cell Tutorial on Facilitated Calcium Diffusion in the Intestinal Epithelial Cell, see the 'simulation' tab.
One may notice the remarkable parallel with the virtual string equation on some of my pages about that subject, except of course that this is 3D simulation (and probably doesn't require hundreds of thousands of CD quality output sound samples), and of course there is no coincidence, wave equations are coupled differential equations of a few orders in time and space.
Biochemistry, fourth edition, Lubert Stryer; Stanford university, 1995, Freeman and Company, New York
Molecular biology of THE CELL, second edition, Alberts, Bray, Lewis, Raff, Roberts, Watson; 1989, Garland Publishing
Ultimate Computing, Stuart Hammeroff
Articles
Directional modulation of visual responses of pretectal neurons by accessory optic neurons in pigeons, Y. Gu, Y. Wang and S. R. Wang, Neuroscience, Volume 104, Issue 1, 10 April 2001; Elsevier Science Ltd.
Persistent sodium channel activity mediates subthreshold membrane potential oscillations and low-threshold spikes in rat entorhinal cortex layer V neurons, N. Agrawal, B. N. Hamam, J. Magistretti, A. Alonso, and D. S. Ragsdale, Neuroscience, Volume 102, Issue 1, 2 Jan 2001, Pages 53-64; Elsevier Science Ltd.